INTRODUCTION
The endocannabinoid system (ECS) is composed of the classic cannabinoid receptors 1 and 2 (CB1R and CB2R, respectively), their endogenous ligands, namely endocannabinoids, and enzymes involved in the synthesis and degradation of these molecules [1,2]. Both cannabinoid receptors (CBRs) are, in fact, part of the heptahelical G protein-coupled receptor (GPCR) family and can inhibit adenyl cyclase activity or even stimulate mitogen-activated protein kinases (MAPKs), being, therefore, classified as Gi/0 receptors [3-5]. CB1R is mainly found in the central and the peripheric nervous system (CNS and PNS, respectively) [6]. In the gastrointestinal tract, this receptor is present in the myenteric plexus and submucous plexus, endocrine cells, and enterocytes [7]. Likewise, CB2R can also be found in the CNS and PNS; however, it is also found in cardiovascular, intestinal, and reproductive systems, as well as bones and adipocytes, being largely expressed in immune cells [8].
Endocannabinoids are the endogenous ligands of the CBRs. They are fatty acid-derived molecules, synthesized by membrane phospholipids [9,10]. Calcium influx works as a synthesis trigger for these compounds in postsynaptic neurons [11], and, as they are released in the synapse, they will act as retrograde messengers, modulating the release of neurotransmitters [12,13]. Besides neurons, several other cells can also synthesize endocannabinoids, such as endothelial and mucosal cells from the intestines or even some immune cells [14-18]. Some of the most well-known and well-described endocannabinoids are N-arachidonoyl-ethanolamine (anandamide or AEA) and 2-arachidonyl-glycerol (2-AG) [19]. AEA is known to be a high-affinity CB1R agonist, with a low affinity for CB2R, being almost inactive in several models, whereas 2-AG is an agonist for both CB1R and CB2R, but with a lower affinity for the latter [20]. Both endocannabinoids will be hydrolyzed inside the cell; after reuptake, AEA is hydrolyzed by fatty acid amide hydrolase (FAAH), and 2-AG by monoacylglycerol lipase (MAGL). Both can also be oxidized by cyclooxygenase-2 (COX-2) and lipoxygenase, generating prostaglandins, ethanolamines, and several other intermediate subproducts [21,22]. Interestingly, the phytocannabinoids, too, such as cannabidiol (CBD), cannabidivarin, and cannabigerol (CBG), might activate or inhibit the CBRs or enzymes involved in the synthesis and degradation of endocannabinoids, modulating the actions of this endogenous system [23].
Activation of CBR1 and CBR2 in the CNS and actions directly in the periphery by the activation of the enteric nervous system (ENS) and resident cells control several intestinal functions, such as intestinal motility, secretion, epithelial permeability, and immunological reactions [24-26]. Both receptors may be involved in the control of gastrointestinal functions during the inflammatory process [27,28]. Thus, patients with colitis had increased levels of AEA and 2-AG that were related to controlling the local inflammation [29-31].
Therefore, several cannabinoids are being studied for the treatment of acute intestinal disorders such as dysmotility, emesis, and abdominal pain and chronic diseases such as irritable bowel syndrome and inflammatory bowel disease (IBD), which by convention is divided into 2 subtypes: Crohn’s disease (CD) and ulcerative colitis (UC) [2].
UC is characterized by inflammation in the mucosa of the large intestine from the anal orifice up to, potentially, the proximal segments. Conversely, CD is characterized by inflammation that can compromise any gastrointestinal tract region, causing ulcers, strictures, abscesses, and fistulas, but in most cases it affects the region close to the ileocecal valve [32]. The cost of IBD diagnosing, hospitalization, and treatment is usually high and treatments are not effective [33,34].
Given this context, this article aims to review the involvement of the cannabinoid system in IBD, CD, and UC and to highlight possible therapeutic approaches for these diseases with the use of cannabinoids.
METHODS
The literature review on the involvement of the cannabinoid system in IBD, with a focus on UC and CD, was carried out using the PubMed database. The descriptors used in the research were “cannabinoid chronic intestinal disease,” “cannabinoid Crohn disease,” “cannabinoid colitis,” “intestinal inflammation,” and “endocannabinoid system.” As inclusion criteria for our review, we selected publications that contained at least one of the descriptors of interest and that were in English. We used no filters for publication date and article type. Therefore, information from full texts, studies in experimental or human models, case reports, meta-analyses, and reviews were included.
ENDOCANNABINOID SYSTEM
Different subtypes of CB1R have been identified in humans and murine models. For instance, the canonical full-length CB1Ra is most commonly found in the brain and skeletal muscles, whereas CB1Rb is mainly expressed in the liver and pancreas [1,2]. Similarly, several isoforms of CB2R also display different expression sites. It was observed by Zhang et al. (2015) [3] that mouse CB2Rb is more abundant in the striatum and the spleen than CB2Ra; their expression can be induced by inflammatory events [4,5].
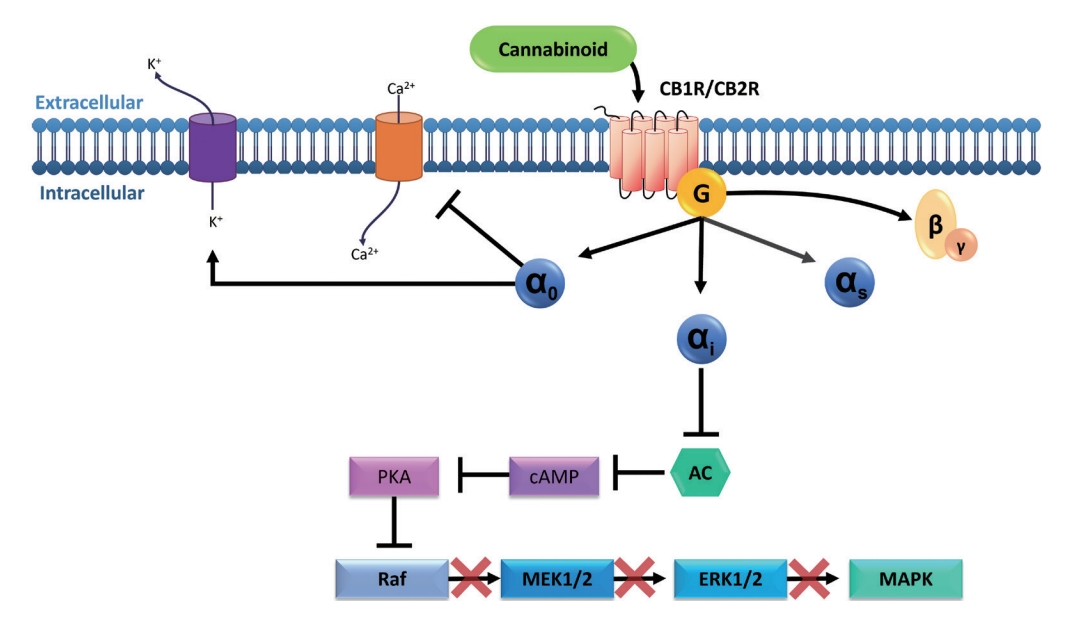
Although both CB1R (a and b) and CB2R (a and b) are members of the GPCR family, it is not unusual to find different signaling events that might be confusing at first sight. This is due to the biased agonism presented by the receptors, meaning that they can adopt multiple conformations leading to different signaling events depending on the stabilization generated by their ligands. In other words, the ligand-biased signal transduction implies that the resulting properties of the agonist or antagonist will depend on the particular signal transduction pathway that is activated (Fig. 1). Therefore, the same molecule might act as an agonist in one pathway and as an antagonist in another [6,7]. These phenomena might explain why the effects of cannabinoid molecules vary among different experimental models and different cell types.
In signaling involving the Gi/0 protein, inhibition of adenylate cyclase (AC) occurs, leading to the consequent inhibition of 3’,5’-cyclic adenosine monophosphate (cAMP) formation and inhibition of protein kinase A (PKA). The effect of PKA inhibition differs depending on cell type [8,9]. In neurons, calcium channels are blocked and potassium channels are opened, leading to hyperpolarization and inhibition of these cells [8]. In intestinal epithelial cells, it is suggested that inhibition of PKA promotes reduced phosphorylation of the cAMP response element-binding (CREB) protein, a transcription factor that regulates cell survival, and consequently a reduction in the rate of cellular apoptosis [10]. In intestinal smooth muscle cells, there is a reduction in the contractility response due to the blockage of calcium channels, which promotes a reduction in actin and myosin interaction [9,11].
Interestingly, in Zebrafish embryos, the activation of CB2R by 2-AG has been shown to inhibit leukocyte migration in response to acute injury [12]. Likewise, another study pointed out that AEA, through the activation of CB2R, promotes the suppression of the release of the inflammatory cytokines interleukin-2 (IL-2), tumor necrosis factor α (TNF-α), and interferon-γ (IFN-γ) from activated human peripheral T lymphocytes [13,14].
The phosphatidylinositol 3-kinase (PI3K) pathway is also stimulated during CB1R and CB2R activation; however, the response depends on stimulus type and microenvironment [2]. When activated, PI3K interacts with insulin receptor substrates, producing phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which will regulate the location and activity of several kinases, including protein kinase B (PKB or Akt). Akt is a downstream target of PI3K and, upon phosphorylation and release into the cytoplasm, will respond to growth factors such as insulin, IL, and epidermal growth factor receptor (EGFR), modulating several cellular processes, such as migration, metabolism, and apoptosis [8,9,15].
Upon activation of the PI3K/Akt pathway, Huang et al. (2011) [15] reported an increase in p-Akt cells in the mucosal and submucosal layers of patients with UC, when compared to normal biopsy tissues. In the same study, the effects of a blocker of the PI3K/Akt pathway were analyzed, and it was found to relieve symptoms, reducing inflammation and TNF-α secretion. Similar results were found by Chen et al. (2017) [16], who described that apoptosis is dysregulated during UC progression, with proapoptotic processes in intestinal epithelial cells and antiapoptotic effect of inflammatory and immune cells.
Another interesting mechanism comprises the p38 MAPK pathway, a critical signaling pathway in bowel inflammation, modulating cellular death, growth, and differentiation [17]. In epithelial cells, MAPK is responsible for proliferative signals by EGFR [18], and p38 MAPK is also found to be increased in CD patients [19]. Furthermore, this pathway could also be involved in cytokine release [17]. However, among the MAPK family members, the extracellular signal-regulated kinase 1 and 2 (ERK1/2), p38 kinase, and Jun N-terminal kinase (JNK) are of the utmost importance in the promotion of apoptotic events, playing an important role in UC pathogenesis; therefore, the inhibition of this pathway led to a reduction in proinflammatory cytokines and cellular infiltrate [20].
In immune cells, the MAPK pathways are involved in the cells’ development, apoptosis, migration, and response against pathogenic stimuli. In microglial cells, CB2R activation through anandamide, an endocannabinoid, has been shown to induce the activation of ERK1/2 and JNK MAPK; this activation resulted in a slight increase in IL-10 levels but also in the inhibition of proinflammatory ILs [21].
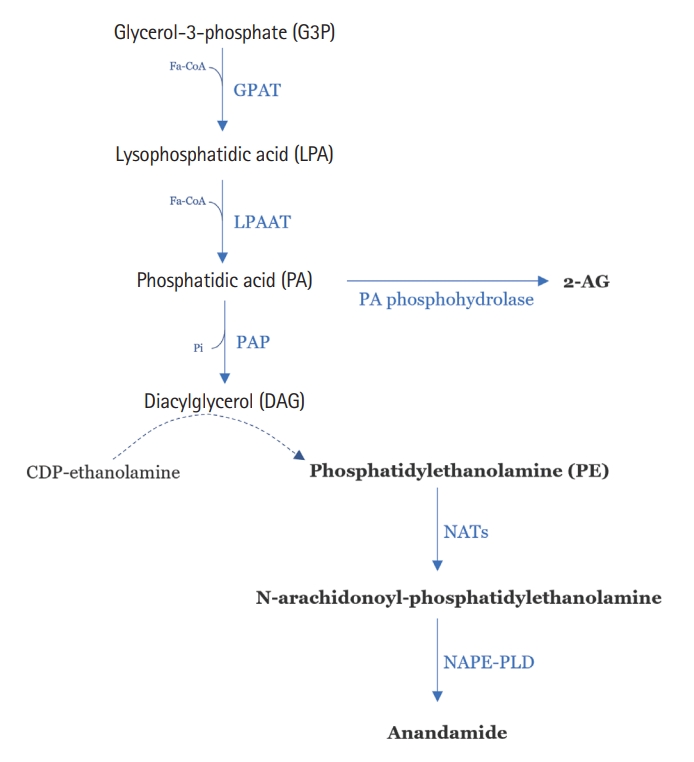
The endocannabinoids are synthesized from membrane lipids in response to specific stimuli, such as neuronal activation or cellular stress, which will promote the influx of calcium into the cell, generating its depolarization [22]. There are at least 15 endogenous compounds that can target CBRs either orthosterically or allosterically. Several of them have been found to bind to cannabinoid CB1Rs in vitro at concentrations in the low or mid nanomolar range [23]. These compounds are anandamide, 2-arachidonoylglycerol, dihomo-γ-linolenylethanolamide, 2-arachidonyl glyceryl ether (noladin ether), docosatetraenoylethanolamide, N-arachidonoyl dopamine, docosahexaenoylethanolamide, and eicosapentaenoyl ethanolamide; the first 3 compounds have also been reported to display a significant affinity for CB2R [23].
N-arachidonoyl-ethanolamine (anandamide or AEA) and 2-AG were the first ones to be discovered and are the most studied until now. The biosynthetic precursors of AEA, the N-arachidonoyl-phosphatidylethanolamines, are produced via N-acyl-transferases (NATs) by N-acyl-phosphatidylethanolamine-selective phospholipase D (NAPE-PLD) or by the sequential action of several enzymes that can also be involved in the synthesis of 2-AG, as reported in Fig. 2 [24]. Both these endocannabinoids will be hydrolyzed inside the cell; after reuptake, AEA is hydrolyzed by FAAH, and 2-AG by MAGL. Both can also be oxidized by COX-2 and lipoxygenase, generating prostaglandins, ethanolamines, and several other intermediate subproducts [25,26]. These lipidic molecules are derived from the acylation of glycerol-3-phosphate (G3P); this molecule will lead to phosphatidic acid (PA) and phosphatidylethanolamine (PE) (Fig. 2) [24,27].
ECS IN THE GASTROINTESTINAL TRACT
The actions of the ECS in the gastrointestinal tract are mainly controlled by the activation of CBRs in the CNS or the periphery, through the ENS or other tissues of the gastrointestinal tract, as shown below [28].
Both CB1R and CB2R, their endogenous agonists, and related enzymes have been detected in the gastrointestinal tract [29]. CB1R and CB2R are widely expressed in intestinal tissue and its innervations [29,30]. CB1R expression is seen in the ENS, including the submucosal plexus and myenteric plexus, and in epithelial and enteroendocrine cells. CB2Rs are expressed in the ENS, also covering the submucosal plexus and myenteric plexus, in epithelial cells in the presence of inflammation, and in immune cells [29].
In the CNS, the regulation of appetite, energy balance, and food intake can be performed by CB1Rs expressed in the lateral hypothalamus, arcuate nucleus, and paraventricular nucleus of the hypothalamus [2,31-33]. Thus, the activation of the ECS in these sites promotes increased appetite, stimulation of lipogenesis, and an increase in energy resources [34]. The control of food intake by the ECS occurs in conjunction with the action of hypothalamic hormones, such as corticotropin-releasing hormone (CRH) in the paraventricular nucleus, the cocaine-amphetamine-regulated transcribed hormone (CART) in the arcuate nucleus, and the melanin-concentrating hormone (MCH) and orexin-A in the lateral hypothalamus, which are well-known modulators of food intake [39]. Hormones produced in the periphery, such as ghrelin, leptin, insulin, and adiponectin, participate in appetite regulation along with the ECS and hypothalamic hormones [32,34]. Studies carried out in animal models have shown that the orexigenic effects are obtained from the stimulation of the ECS in the CNS and the neurons that release MCH, in addition to the increase in neuropeptide Y (NPY) and the hormones orexin-A and ghrelin [40-43].
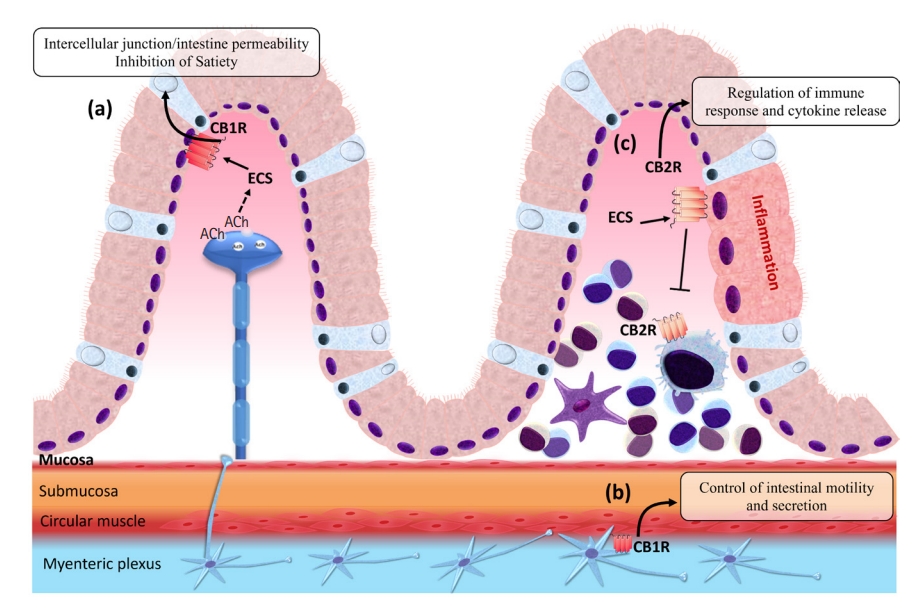
Thus, appetite control also takes place in the small intestine, where during the fasting period or in the presence of taste of dietary fats, endocannabinoid signaling is triggered by the activation of CB1Rs, aiming to inhibit satiety [44,45]. Inhibition of muscarinic receptor type 3 (M3) blocks the induced production of 2-AG in the jejunum. This can be replicated by peripheric administration of CB1R antagonists, confirming that cholinergic signaling, possibly through the vagus nerve, activates the muscarinic receptor of acetylcholine (mAChR) in the jejunum, stimulating the conversion of 1-stearoyl-2-arachidonoyl-sn-glycerol (SAG) into 2-AG by the enzyme diacylglycerol lipase (DAGL), stimulating food intake (Fig. 3, part a) [45,46]. Besides these mechanisms for controlling appetite and food intake, there are also CB1Rs expressed in the enteroendocrine cells of the small intestine, which, when activated, promote hunger during the fasting period and boost the intake of high-fat foods, inhibiting the release of cholecystokinin (CCK) that would normally bind to CCK receptors inducing satiety after a meal [44,45]. The control of intestinal motility, secretion, epithelial permeability, and immunology are also accomplished by the modulation of CB1R and CB2R expressed in the ENS and resident cells [47,48].
The activation of CB1R in the ENS is mainly related to the negative control of cholinergic neurotransmission, with consequent inhibition of the action of this neurotransmitter on muscarinic receptors and a reduction in intestinal smooth muscle contractions (Fig. 3, part b) [49-51]. Activation of CB2R by the agonist JWH-133 in rats promotes a reduction in the speed of food passage through the gastrointestinal tract, as a consequence of the decrease in muscle contractions [52]. Thus, the activation of this receptor in the ENS, as well as the activation of CB1R, promotes the inhibition of the contractility of the intestinal muscles, inhibiting the release of ACh [53].
Activation of CB1R and CB2R promotes the inhibition of proinflammatory cytokine secretion (Fig. 3, part c), suppression of macrophage and mast cell activation, modulation of TH1 and TH2 lymphocytes through induction of apoptosis, and inhibition of proliferation and consequently reduced inflammation [56-59]. Di Marzo and Izzo (2006) [60] discussed that the ECS is overstimulated in the intestines and colon during inflammatory disorders. Enhanced production of endocannabinoids is part of the gastroprotective mechanism and several other regulatory functions-including motility and secretion-that might function through CB1R [29,61]. CB2R is either absent or only weakly expressed in the human intestinal epithelium; however, in IBD it becomes evident in the luminal membranes at ulcerative margins, and immune cells in the lamina propria (LP) might also express CB2R [29,62]. Therefore, differently from CB1R, CB2R appears to be less expressed during homeostasis, but it is inducible during inflammatory disorders [21].
There is evidence that CB1R and CB2R are also related to the control of sensations such as visceral pain, emesis, and nausea, which are sensory experiences that may be linked to the functioning of the gastrointestinal tract [29]. Sanson et al. [63] demonstrated that CB1R and CB2R agonists attenuated the basal sensitivity and that this was increased by colon inflammation in rats. Moreover, the intestinal microbiota is known to contribute to several improvements in digestive health. It was demonstrated that the probiotic agent Lactobacillus acidophilus promoted an increase in the expression of CB2R in the epithelium of rats and mice and a consequent reduction in visceral sensitivity in cases of irritation and inflammation [64]. A possible mechanism for pain regulation from CB2R would be the inhibition of the release of inflammatory mediators from the epithelium that would activate the primary afferent nerves and cause visceral pain [65]. Furthermore, cannabinoids are effective antiemetics and have been used in the clinic for years; however, they are not the first-line therapeutic agents [66]. Some evidence also demonstrates the involvement of CB1R in the brainstem [67,68] and that of CB2R in the dorsal vagal complex for the regulation of emesis [69]. Additionally, in animal models, activation of the ECS in the insular cortex has been found to regulate nausea, and when endocannabinoid levels were increased by administering an FAAH or MAGL inhibitor, nausea was reduced. However, it is suggested that only MAGL inhibition is enough to generate the effect against nausea by the activation of CB1R by 2-AG, and AEA seems not to be involved in this event [70-72].
Therefore, the activation of CB1R and CB2R promotes a reduction in colonic motility; control of the secretion of neurotransmitters such as acetylcholine and hormones such as ghrelin, leptin, adiponectin, and insulin; control of the permeability of the intestinal epithelium; and regulation of visceral pain, emesis, and nausea, besides anti-inflammatory and antiproliferative effects [73,74]. Considering the various functions performed by the ECS in the gastrointestinal tract, changes in its tone can lead to several effects, which can be explored for the development of drugs for the treatment of various diseases involving disorders of the gastrointestinal tract.
ECS IMPACT IN CHRONIC IBDs
IBD is a destructive inflammatory disorder and comprises both UC and CD. Therefore, UC is an intestinal inflammatory disorder of the colon, characterized by the infiltration of immune cells and the release of inflammatory mediators that may lead to ulcers and disruptions in the mucosa [75,76]. Likewise, CD is a chronic inflammatory condition of the gastrointestinal tract, resulting from a complex interplay of factors, such as genetic susceptibility, gut microbiota, and environmental pressures, leading to dysregulations in the patient’s immune response [77]. However, UC is a confluent mucosal inflammation, being initiated at the anal verge and spreading proximally for a variable length. CD can affect any region of the gastrointestinal tract, resulting in ulcers with transmural inflammation leading to fibrosis, fistula, and strictures, usually close to the ileocecal valve. Additionally, the dispersed inflammation in CD can leave areas of normal mucosa in the midst of inflammation, called “skip areas.” [9]
IBD is a chronic disease that can reduce the life quality of patients and is correlated with extraintestinal comorbidities, such as cardiovascular diseases, osteoporosis, arthritis, and lymphomas [9,78-80]. The symptoms of IBD can be further exacerbated by stress and psychiatric diseases, such as depression and anxiety [81].
Corticosteroids and immunomodulatory drugs, which are the basis for the treatment of IBD, do not always provide satisfactory outcomes [75,82,83]. As demonstrated, the ECS is largely expressed in the gastrointestinal tract; therefore, the manipulation of this system becomes an interesting approach to be investigated.
CHANGES IN THE ECS IN UC
The expression and activation of CBR in experimental models of intestinal inflammation have been investigated by several researchers. Engel et al. (2010) [84] found that CB1R (-/-), CB2R (-/-), and CB1R/CB2R (-/-) knockout C57BL/6 mice were more susceptible to the development of UC induced by 2,4,6-trinitrobenzenesulfonic acid (TNBS). Storr et al. (2009) [85] also evaluated the participation of the ECS in an experimental model of TNBS-induced colitis in wild C57BL/6 mice and CB2R (-/-) knockout mice, and these mice were treated with the agonists JWH-133 and AM-1241 and the antagonist AM630, selective for CB2R. CB2R (-/-) knockout mice had increased inflammation and did not respond to agonist or antagonist treatments. In wild-type (WT) mice, treatment with the agonists JWH-133 and AM-1241 was found to reduce intestinal inflammation. In contrast, the use of the AM-630 antagonist in WT mice exacerbated these parameters, demonstrating an increase in inflammation. Furthermore, an increase in CB2R mRNA expression was found during inflammation in WT mice. Another study with CB2R (-/-) knockout C57BL/6 mice demonstrated that these animals have worsened colitis induced by dextran sulfate sodium (DSS) compared to WT mice [86].
Furthermore, changes in endocannabinoid metabolizing enzymes have been reported in female C57BL/6N mice, genetically modified to express less FAAH. It was shown that this enzyme deficiency generates protection against dinitrobenzene sulfonic acid (DNBS)-induced UC, as revealed by a reduction in macroscopic and microscopic damage and in myeloperoxidase (MPO) enzyme activity [61]. Storr et al. (2008) [87] demonstrated that FAAH mRNA is increased on the 3rd day after colitis induction with TNBS in mice. Recall that FAAH is the main enzyme responsible for AEA hydrolysis, so an increase in this enzyme might reduce the endocannabinoid concentration during the inflammatory process. Likewise, Shamran et al. (2017) [88] showed that FAAH inhibition ameliorates the clinical score and reduces the frequencies of activated CD4+ T cells in the colon and proximal lymphoid tissues. Furthermore, Izzo and Sharkey (2010) [89] have demonstrated that mice without or with reduced FAAH have elevated basal levels of AEA and are more resistant to TNBS-induced colitis than wild mice. Borrelli et al. (2009) [90] used male Institute of Cancer Research (ICR) mice strain and found that during DNBS-induced inflammation, the concentrations of AEA and 2-AG increased, and the mRNA expression of the FAAH enzyme in the colon decreased, revealing the physiological function of the ECS. Likewise, Tourteau et al. (2014) [91] used substances derived from 3-carboxamide-5-arylisoxazole CB2R agonists and FAAH inhibitors in the treatment of DSS-induced colitis in mice and showed significant improvement in inflammation. In another study, the use of an FAAH inhibitor, PF 3845, reduced TNBS-induced experimental UC due to increased endocannabinoid expression and activity [92]. Furthermore, a study using ARN2508, an inhibitor of FAAH, demonstrated an improvement in inflammation, a reduction in the mortality rate, and a decline in weight loss in mice with TNBS-and DSS-induced colitis. In this case, as mentioned above, the anti-inflammatory effect obtained by inhibiting FAAH is probably related to an increase in endocannabinoids such as AEA [93].
Regarding MAGL activity, in C57BL/6N mice with DSS-induced colitis, real-time polymerase chain reaction (RT-PCR) analysis demonstrated that the expression of genes encoding this enzyme was reduced during colon inflammation [94]. In addition, in mice with TNBS-induced colitis, inhibition of the MAGL enzyme by the selective inhibitor JZL184 promoted an increase in 2-AG, which was associated with a reduction in macroscopic and histological damage of the colon, as well as a decrease in the expression of inflammatory cytokines [95].
In humans, Western blot analysis and immunocytochemistry in colon tissue demonstrated an increased expression of CB2R, the enzyme DAGL, and MAGL in the colonic epithelium, mainly in patients with mild and moderate colitis. In contrast, in patients with inactive colitis, the expression of CB1R and CB2R and the DAGL enzyme was reduced, but the expression of the NAPE-PLD enzyme was increased, especially in patients treated with corticosteroids [96]. Wright et al. (2005) [62] also found an increased expression of CB2R in the human colonic epithelium during UC, demonstrating an immunomodulatory function of the ECS.
Besides the role of enzymes in the physiopathology of IBD, the endocannabinoid AEA appears to be the main modulator of colon-induced inflammation. AEA was found to be increased in colon biopsy from patients with untreated UC, as analyzed by liquid chromatography with detection by mass spectrometry [97]. Harvey et al. (2013) [98] showed that the incubation of human colon tissues with the inflammatory cytokines TNF-α and IL-1B causes damage like UC. Administration of AEA and a selective CB2R agonist, JWH-015, showed a significant improvement in inflammation, as evidenced by a reduction in mucosal crypt loss, luminal epithelial damage, and lymphocyte infiltration in the LP. These effects were partially reversed by the administration of a CB2R inverse agonist, JTE-907, which demonstrates that the anti-inflammatory effects of JWH-015 and AEA are dependent on the activation of this receptor [99].
Grill et al. (2019) [100] evaluated the regulation of the ECS in 17 patients diagnosed with UC and found an increase in AEA expression and a decrease in mRNA expression of the NAPE-PLD enzyme, which seems contradictory, considering that NAPE-PLD participates in the synthesis of this endocannabinoid. However, to explain this fact, the authors suggested that there is an allosteric feedback inhibition of NAPE-PLD by AEA or a reduced degradation of the endocannabinoid lipids. Furthermore, other enzymes besides NAPE-PLD may be responsible for the synthesis of AEA. In another study by Suárez et al. (2012) [101], human biopsy from healthy patients with untreated active colitis and from patients treated with 5-aminosalicylic acid (5-ASA), glucocorticoids, and/or immunomodulators until disease remission were analyzed using RT-PCR and immunohistochemistry to assess the expression and distribution of FAAH and NAPE-PLD in the intestine. RT-PCR analysis of the mucosa from untreated UC individuals revealed an increase in the gene expression of FAAH and no alterations in the gene expression of NAPE-PLD. Also, with immunohistochemical analysis of the colon epithelium from untreated UC patients, the authors observed a decrease in NAPE-PLD and the conservation of FAAH levels, as compared to healthy individuals. However, after treatment with 5-ASA and glucocorticoids, an increase was found in the expression of NAPE-PLD in the epithelium, indicating a role of this enzyme in UC remission [101].
Besides the above-described studies, there are several studies showing the relevancy of the ECS in the modulation of inflammation during UC by increasing the expression of receptors, by increasing the synthesis of endocannabinoids, or by inhibiting or activating the enzymes involved in the degradation or synthesis of endocannabinoids. In summary, changes in the expression of the ECS components during UC can generate anti-inflammatory effects in animal and human models, as described in Table 1.
CHANGES IN THE ECS IN CD
As in UC as discussed above, it has been suggested that in CD the ECS may act to reduce the local inflammatory response. Therefore, it has been the topic of several studies to test and evaluate changes in the expression of CBR and the synthesis and degradation of endocannabinoids during this intestinal inflammation. The single nucleotide polymorphism 1359 G/A of the CB1R gene has been shown to influence the CD phenotype. Individuals who were homozygous for the G allele showed a higher probability of developing CD before the age of 40 years, demonstrating that CB1R influences the manifestation of CD [103]. The expression of CB2R and AEA was evaluated in ileum tissue of mice and in CD4+ T cells of an experimental model of chronic ileitis. The results demonstrated an increased expression of CB2R and AEA in mice with inflammation compared to control mice. Moreover, an 11-fold increase in CB2R mRNA expression was found in regulatory T cells, demonstrating the important role of the ECS in the regulation of inflammatory response [104].
The expression of CB2R was also analyzed in histological sections of biopsy samples of the intestine of patients with CD. There was no difference in the expression of this receptor between CD patients and healthy individuals. However, the expression of CB1R was shown to be significantly increased in patients with CD [105]. Moreover, Grill et al. (2019) [100] found that the concentrations of AEA and 2-AG were markedly increased in the plasma of CD patients. Furthermore, in the intestinal mucosa, an increased gene expression of synthesizing (NAPE-PLD and DAGL) and metabolizing (FAAH and MAGL) enzymes and a decreased transcription of CB1R were identified [106].
On the other hand, Di Sabatino et al. (2011) [59] found decreased levels of AEA in the inflamed mucosa of patients with IBD compared to non-inflamed tissue, which was concomitant with lower activity of the NAPE-PLD enzyme that synthesizes this endocannabinoid and higher activity of the FAAH enzyme that degrades this ligand. Some divergences were found between these studies, and those may be related to the different tissues evaluated and to the change in the expression of the ECS according to the stage of the disease [107].
Therefore, the physiological or induced changes in the ECS during CD that can generate an anti-inflammatory effect in animal models and humans are summarized in Table 2.
CANNABINOIDS AS POSSIBLE DRUGS FOR THE TREATMENT OF CHRONIC IBDs
Given the various disadvantages and limitations of the treatments available for IBD, it is relevant to develop new pharmacological alternatives for the treatment of UC and CD. In this context, cannabinoids can be a good pharmacological strategy for the treatment of IBD because of the various functions of the ECS in maintaining intestinal homeostasis and the anti-inflammatory effects observed by the positive regulation of this system in animal and human models.
Phytocannabinoids such as CBD, Δ9-tetrahydrocannabinol (THC), CBG, cannabinol (CBN), and cannabichromene (CBC), all isolated from Cannabis sativa, popularly known as marijuana, were evaluated as possible drugs for the treatment of UC and CD. However, when using some of these substances, with the clear exception of CBD, it is necessary to pay attention to the possibility of dose-dependent psychoactive effects [119].
CBD administration was evaluated in rats and mice with intestinal inflammation induced by irritating substances, such as croton oil (CO), lipopolysaccharide (LPS), and DNBS. It was observed that CBD reduced edema, hyperemia, adhesions in the small intestine, inflammatory cell infiltration, colon weight/length ratio, and intestinal hypermotility [90,108-110]. Additionally, there was a decrease in inflammatory cytokines, such as IL-1β, and an increase in anti-inflammatory cytokines, such as IL-10 and IL-6 [90,109]. Other studies have found decreased MPO activity, reduced induced nitric oxide synthase (iNOS) expression, and reduced nitric oxide (NO) synthesis and its stable metabolites [90,110,111]. Therefore, there is evidence that the use of CBD improves intestinal inflammation in experimental models. Similarly, the administration of CBD-rich C. sativa extract in mice promoted a reduction in intestinal motility, colon weight/length ratio, and MPO activity, indicating an improvement in inflammation [113].
However, Naftali et al. (2017) [112] evaluated the administration of CBD in humans with CD and found no significant improvement in intestinal inflammation. Therefore, there is a need for more studies involving CBD in humans with CD. In contrast, the administration of CBD-rich C. sativa extract in humans with UC demonstrated greater remission of the disease, better patients’ quality of life, and a reduction in the total Mayo score, which is used to assess the UC stage by scoring the pattern and the presence of bleeding in the stools, endoscopic results, and medical evaluation [114].
THC is a cannabinoid present at high concentrations in C. sativa; it has psychoactive activity and is therefore of limited use [119]. Conversely, in some studies, the use of this phytocannabinoid presented beneficial effects on intestinal inflammation when administered alone or in conjunction with other cannabinoids.
Jamontt et al. (2010) [115] demonstrated that the administration of THC alone or in combination with CBD resulted in decreased weight gain and macroscopic damage in the intestines of rats, in addition to reducing MPO activities and improving the contractile response of the intestinal muscles. In humans with CD, the administration of THC promoted an increase in the patients’ quality of life, greater remission of the disease, and improved appetite and sleep [116]. Regarding the simultaneous use of CBD and THC, it is worth mentioning that some studies report that the negative allosteric modulation of CBD in CB1R might reduce the adverse effects of THC [128].
Another phytocannabinoid with potential application in the treatment of IBD is CBN, a product of THC oxidation, found in large quantities in C. sativa [129]. In a model of intestinal inflammation using CO, it was found that CBN inhibited the increase in intestinal motility and contributed to an anti-inflammatory effect [117]. Nonetheless, further studies are required to better understand and elucidate the applications of CBN.
Besides these compounds, another potential drug for the treatment of IBD is CBG, which, like CBD, does not display psychotropic effects [119,130,131]. Borrelli et al. (2013) [118] analyzed the effect of CBG administration in male ICR mice, in peritoneal macrophages, and in epithelial cells of the PTK6 knockout colon of these animals. In mice that received this drug, it was found that there was a reduction in the weight/length ratio of the colon, damage to the colon, intestinal permeability, MPO activity, iNOS expression, and levels of IL-1β and IFN-γ. The authors also described an increased superoxide dismutase (SOD) activity and IL-10 levels. Finally, Ki-67 cells were detected in the lower half of the mucosa, thus indicating their proliferation. In mice untreated with CBG, Ki-67 was detected mainly in inflammatory cells. In the peritoneal macrophages, the synthesis of nitrites was decreased, and the levels of iNOS were reduced. In the epithelial cells of the PTK6 knockout colon, the synthesis of reactive oxygen species (ROS) was reduced. Therefore, the authors suggested an anti-inflammatory effect of CBG in the colon.
Likewise, another phytocannabinoid, CBC, has shown a protective effect against IBD. In mice with CO-induced inflammation, it was found that the administration of CBC promoted a decrease in intestinal motility and inhibited ileum contractions induced by ACh and electrical field stimulation (EFS) [99]. In another study, in mice with DNBS-induced inflammation, the administration of CBC led to a reduction in tissue damage, the colon weight/length ratio, intestinal permeability, and MPO activity. Furthermore, CBC reduced the effect of DNBS on cell proliferation, with mitotic activity being restricted to half of the mucosa, as identified by Ki-67 immunoreactivity [106].
In Table 3, we summarize the studies that investigated the applications of phytocannabinoids as potential treatments for intestinal inflammatory disorders.
Synthetic cannabinoids, such as the non-selective CBR agonists AM-841, HU-210, and WIN55,212-2, the CB1R agonists arachidonyl-2’-chloroethylamide (ACEA) and CP55,940, and the CB2R agonists AM-1241, β-caryophyllene, compound 26, HU-308, and JHW-133, are analogs of endocannabinoids and phytocannabinoids or compounds that have been synthesized to mimic the action of natural cannabinoids [134].
Studies in mice with induced intestinal inflammation by irritating substances, such as DSS, DNBS, and TNBS, in which the mice were treated with non-selective CBR agonists (AM-841, HU-210, and WIN55 212-2), mainly showed that the administration of these drugs generated protection against intestinal inflammation, as evidenced by decreased macroscopic and microscopic damage, inflammatory infiltrate in the intestine, and MPO activity [61,120-123].
Similarly, the administration of synthetic CB1R agonists (ACEA and CP55,940) in mice with induced intestinal inflammation has shown that these substances have anti-inflammatory and antioxidant actions. For instance, treatment with ACEA led to weight gain, shortening of the colon, and reduced macroscopic damage, diarrhea, intestinal hypermotility, MPO activity, inflammatory infiltrate, and nitrite production, demonstrating the protective effect of this drug [102,106,124]. The use of CP55,940 demonstrated a reduction in intestinal motility and inflammation [117].
CB2R agonists (AM-1241, β-caryophyllene, compound 26, HU-308, JHW-133) were also evaluated in experimental models of induced inflammation. Macroscopic damage, such as ulcers and adhesions, and microscopic damage, such as thickening of the intestinal wall, shortening of the colon, inflammatory infiltrate, the expression of inflammatory mediators, and the activities of MPO and N-acetylglucosaminidase (NAG), were reduced with the administration of these drugs in mice with induced intestinal inflammation [85,86,125,126].
Furthermore, Bento et al. (2011) [125] observed that the administration of β-caryophyllene promoted a loss in the signaling pathways involving NF-κβ, CREB, ERK1/2, and IKK (as induced by MAPK and transcription factors) and an inhibition of apoptosis and proliferation in colon tissue (as demonstrated by a reduction in the levels of caspase-3 and Ki-67). Another study showed that the administration of HU-308 reduced Nod-like receptor Pyrin Domain Containing 3 (NLRP3), the ratio casp-1 p20/casp-1 p45, and the expression of proIL-1β and IL-1β mRNA (of SQSTM1 in the colon) and increased the levels of LC3-II, LC3-I, and beclin-1, which are related to increased autophagy [86].
Among the selective CB2R agonists, the synthetic drug JHW-133 is one of the most studied in the literature. The administration of this drug in mice and rats with induced intestinal inflammation resulted in a reduction in intestinal motility, weight gain, shortening of the colon, macroscopic damage, diarrhea, MPO activity, inflammatory infiltrate, bodyweight loss, and nitrite levels, leading to an improvement in the inflammatory condition of the animals [52,85,102,106,127].
Furthermore, some authors found that in CB2R-deficient mice or WT mice treated with an antagonist of this receptor, the anti-inflammatory effects promoted by AM-1241, β-caryophyllene, and JHW-133 were not observed, confirming the action of these substances through the activation of CB2R [85,125].
Table 4 shows the description of the animals, the substances used to induce intestinal inflammation, and the results obtained in each of the studies after the administration of CBR agonists (selective and non-selective).
ENZYME INHIBITORS AND CELL REUPTAKE INHIBITORS OF ENDOCANNABINOIDS, AS POTENTIAL DRUGS FOR THE TREATMENT OF CHRONIC IBDs
Several studies have shown that during intestinal inflammation, CBR is overexpressed, endocannabinoid synthesis is increased, and the activity of the metabolizing enzymes of these ligands is reduced to restore the balance in the body [84,86,92,93]. Thus, substances that inhibit the degradation of the main endocannabinoids—AEA and 2-AG by the FAAH and MAGL enzymes, respectively—and compounds capable of inhibiting the reuptake of these endocannabinoids were seen as potential drugs for the treatment of IBD.
The administration of the AEA enzyme hydrolysis inhibitor, arachidonoyl serotonin (AA-5-HT), in mice with DNBS-induced colitis promoted a reduction in macroscopic and histological scores and MPO activity [97]. The use of VDM-11, an AEA reuptake inhibitor, mainly promoted a decrease in macroscopic and microscopic damage, histological score, and MPO activity [87,97].
Moreover, the use of FAAH inhibitors (URB-597, compound 39, and PF-3845) in mice with induced intestinal inflammation demonstrated, above all, a reduction in macroscopic and microscopic damage, histological scores, bodyweight loss, MPO activity, inflammatory cytokines, and mortality rate and even normalized the architecture of intestinal tissues [87,92,132,133]. Furthermore, the administration of ARN-2508, an inhibitor of FAAH, COX-1, and COX-2, demonstrated that the bodyweight loss, mortality rate, and intestinal inflammation of the mice were reduced [93].
In another perspective, JZL-184, a selective MAGL inhibitor, when administered to mice with TNBS-induced intestinal inflammation, reduced macroscopic lesions, decreased submucosal edema, normalized mucosal structure, and reduced leukocyte infiltrates, the levels of the inflammatory cytokines MCP-1 and MIP-1, and the plasma levels of LPS [95].
Thus, several studies in experimental models have shown that the use of endocannabinoid-enzyme inhibitors and endocannabinoid reuptake inhibitors is capable of promoting improvement in the intestinal inflammatory profile, being potential drugs for the treatment of IBD (Table 5).
CONCLUSIONS
As summarized in this review, during UC and CD, the expression of CB1R and CB2R and the synthesis of endocannabinoids are increased, and the activity of cannabinoid metabolizing enzymes is decreased. Thus, potential therapeutic approaches for UC and CD have been identified with the use of CBR agonists, which include endocannabinoids, phytocannabinoids, synthetic cannabinoids, inhibitors of endocannabinoid metabolizing enzymes, and AEA reuptake inhibitors. These compounds, when evaluated in experimental models, demonstrated antiproliferative, antinociceptive, antioxidant, and anti-inflammatory actions, by regulating the levels of inflammatory cytokines, iNOS, NO, ROS, and the activities of the enzymes MPO, SOD, and NAG. In addition, they lead to an increase in the expression of anti-inflammatory cytokines, such as IL-10 and IL-4 (Fig. 4).
Studies in humans are scarce and, when performed, cover a limited sample, making it difficult to assess the efficacy of cannabinoids used in the treatment of UC and CD. Nevertheless, CBR agonists, inhibitors of the reuptake of AEA and 2-AG, and inhibitors of the enzymes that degrade endocannabinoids are promising drugs in the management of these diseases. However, comparative research on these substances is lacking, although such research is necessary to better evaluate the best therapeutic options for long-term use in the treatment of IBD.
Given the importance of the cannabinoid system in gastro-intestinal tract homeostasis (Fig. 5) future clinical studies are needed to better evaluate the risks and benefits of prolonged treatment using substances that interact with this system, the possible drug interactions, and the adverse effects of using cannabinoids, to guarantee the effectiveness of treatment and patient safety.